
Page 1
人体由50万亿到70万亿个细胞组成,起始的单细胞被称为合子(zygote),全能。
Page 2
“见别人已见,但想他人未想”。
Page 3
John Gurdon通过对非洲爪蟾(Xenopus laevis)进行体细胞核移植(somatic cell nuclear transfer, SCNT, 将成年蟾蜍的细胞核从细胞中取出并注入未受精的卵子),用证伪的方式证明了已分化细胞的细胞核在适宜的条件下能够形成一个完整的动物,即遗传物质不会被永久地丢失或失活,只会被表观修饰。
Page 11
聪明的傻瓜经常把事情搞得又大又复杂……而相反的工作则需要一点天分和巨大的勇气。——Albert Einstein
Page 13
卵子的细胞质中的有些东西能够激活被研究者注射到卵子里面的成体细胞核进行重新编程。
Page 14
内细胞群(ICM)需要在实验室里小心培养,但为我们提供了ES细胞(胚胎干细胞,多能)。ES细胞可以形成胚胎和成熟个体具有的所有细胞。而iPS则是通过基因注入成细胞而被诱导而成的多能性干细胞。
Page 18
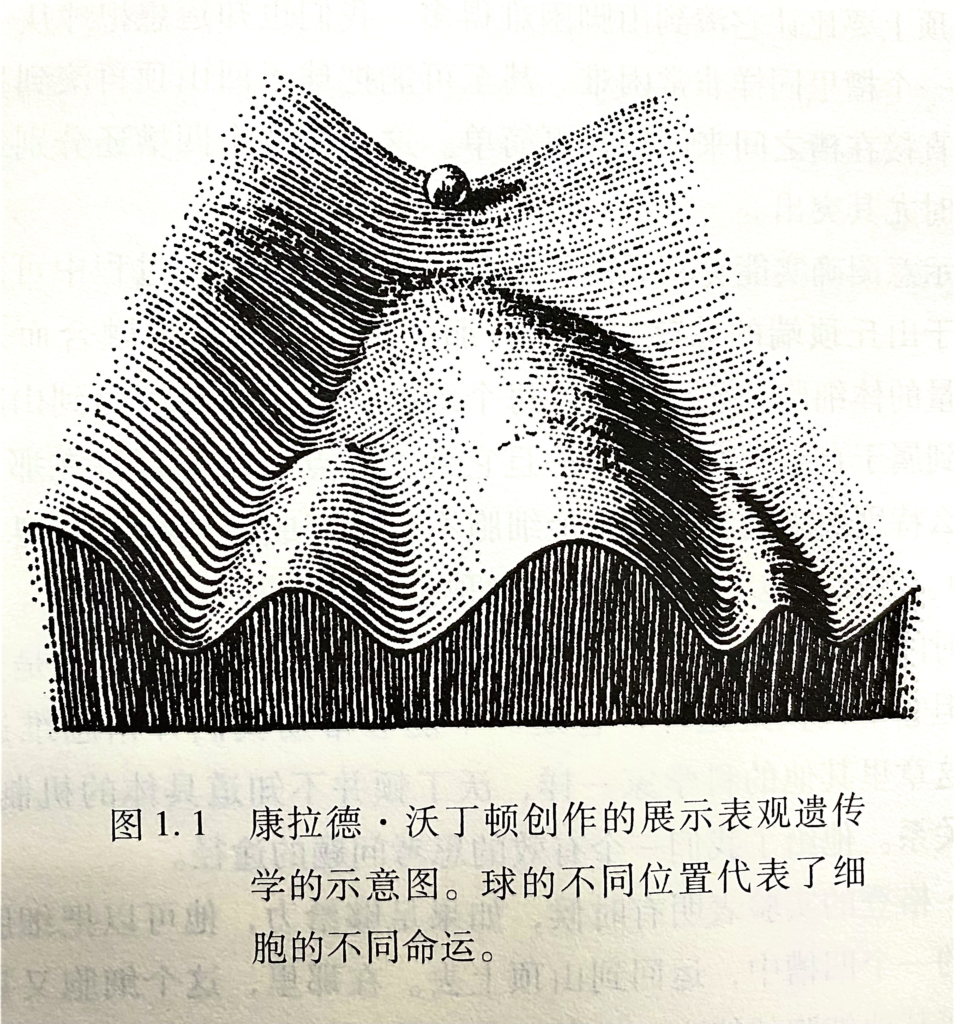
想想看,哺乳动物细胞有大约20000个基因,而只要4个基因(Oct4, Sox2 Klf4, c-Myc)就可以把它变成完全不同的具有多能性的细胞,这是多么不可思议呀。仅靠这4个基因,山中教授就把球从沃丁顿(Waddington’s epigenetic landscape)的山底推上了山顶。
Page 23
山中伸弥(Kazutoshi)制造iPS细胞的实验中,能够重新编成的细胞只有很小的一部分,即使在最好的结果里也不到1%。而让这些iPs细胞获得重新编程要养几周的时间。尽管后续的工作提高了成功的比例而且需要的时间也有所缩短,但仍与自然生殖效率无法相比。为什么?答案就在表观遗传学。当卵子跟精子融合后,这个重新编程过程就高效地进行着并在36小时内完成,因为卵子的细胞质在移除表观标记方面具有极高的效率,就像一个巨大的分子橡皮。
Page 25
诗人能宽容一切,除了拼写错误。——Oscar Wilde (The children of the Poets)
Page 26
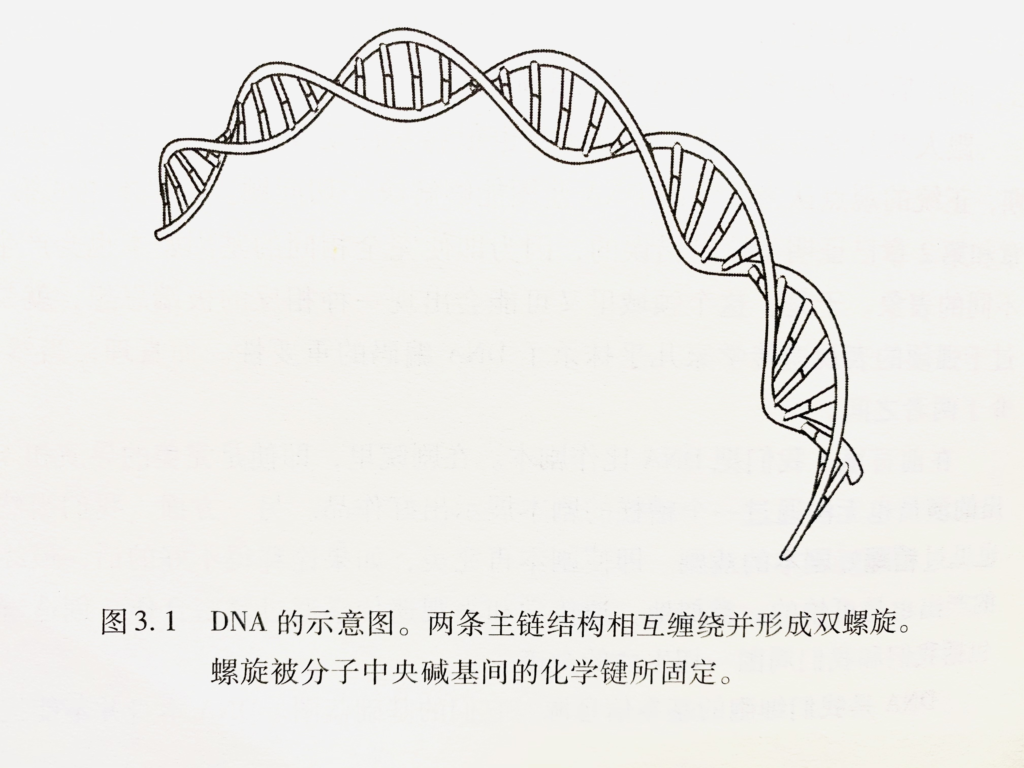
DNA(Deoxyribo nucleic acid)是携带所有蛋白质编码的载体。人类是如此复杂,而DNA只是一种由4个字母组成的语言。ACGT。C是表观遗传学里最重要的碱基。
Page 29
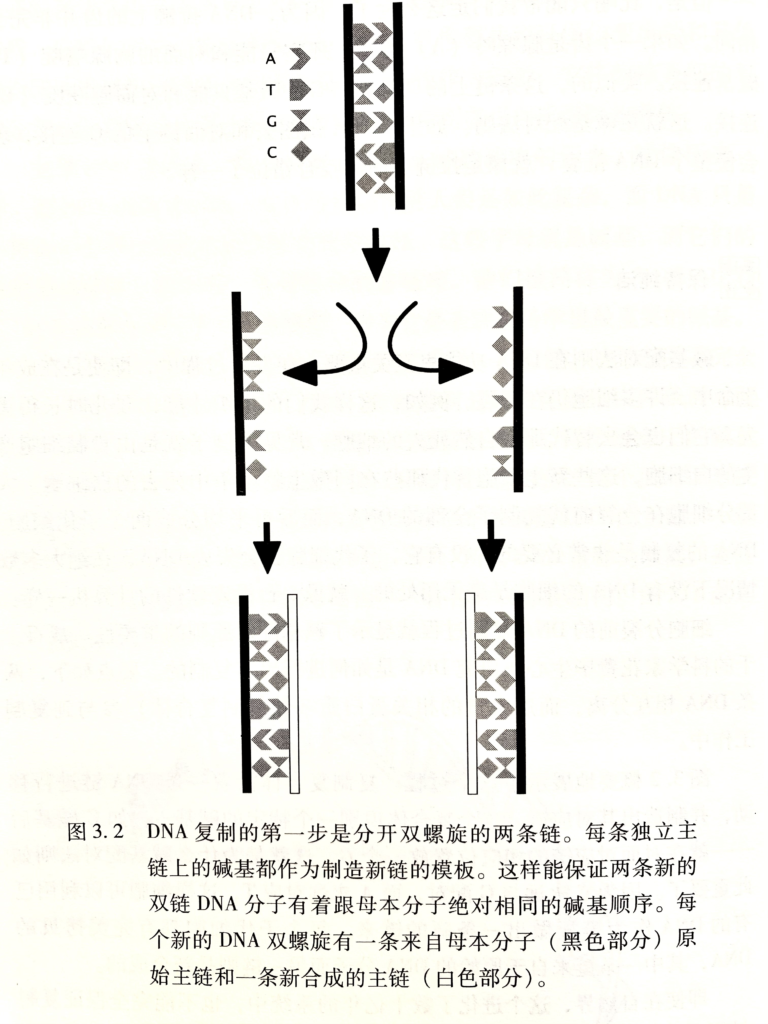
随着时间不停地过去,人的身体里会有大约50,000,000,000,000个具有完全一致的DNA的细胞。当我们知道每次从一个细胞分裂成两个子代细胞需要复制多少个DNA的时候,也许会有些吃惊。每个细胞里有60亿个碱基对(一半来自父亲,一半来自母亲)。这60亿个碱基对的顺序被称为基因组。所以,身体里面每个细胞都是复制这60亿个碳基的产品。
Page 30
蛋白质跟乐高类似,蛋白质中的“积木”是相当小的分子,叫氨基酸,在蛋白质的乐高玩具中一共有20种标准氨基酸作为积木。但是这20种标准氨基酸通过不同的排列顺序和长度的组合,能够创建出一个令人难以置信的数量庞大的蛋白质群。
Page 30
但是还有一个问题,即使氨基酸的数量只有20种,那这20种氨基酸又是如何被DNA中的4个碱基对编码的呢?解决方案就是细胞“阅读”DNA的方式是通过每次包括3个碱基的阅读框实现的。每个包括3个碱基的阅读框被称为密码子,ACGT这4个碱基能够产生64(4^3)种不同的组合方式,对于编码20个氨基酸来说已经绰绰有余了。例如,由AAA和AAG编码的氨基酸叫作赖基酸。
Page 30
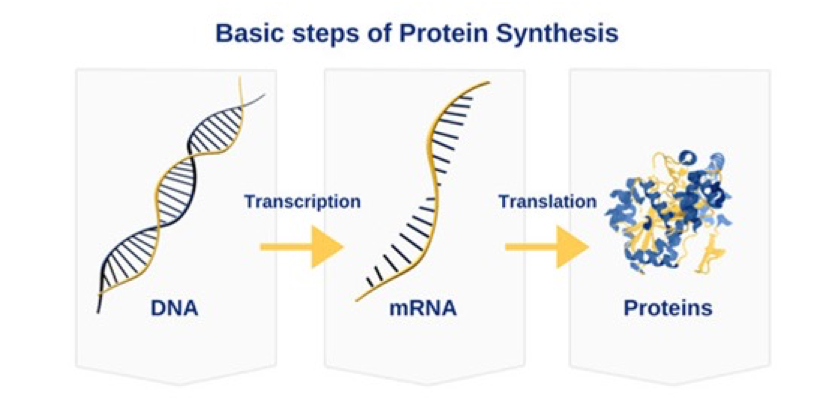
染色体内的DNA作为脚本生成蛋白质需要一个中介,一个叫做信使RNA(mRNA)的分子。mRNA与DNA很像,但有些细节不同。它的主链与DNA稍有不同(所以叫RNA,因为它由Ribonucleic acid 核糖核酸,而不是脱氧核糖核酸组成);它是单链结构(只有一条主链);它的T被结构近似但稍有区别的U取代。当特定的DNA准备被“阅读”以产生蛋白质时,一个巨大的蛋白质复合体将DNA上适当位置的双链打开,并生成mRNA拷贝。这个复合体利用碳基配对原则制作出完美的mRNA拷贝。随后mRNA分子被作为临时模版在细胞内一个特殊的结构中生产出蛋白质。这个特殊的结构通过阅读“三字密码”而将相应的氨基酸拼接起来形成蛋白质长链。
Page 31
进化选择了这种方式,那就一定有很多理由。其中之一就是防止对原始文档DNA的损害。当DNA解开双螺旋时,就处于易伤状态,而细胞在进化中会尽量避免其发生。DNA编码蛋白质的这种间接方式能够将DNA开放且易伤的时间期限最小化。还有一个原因就是这样可以对特定蛋白质的表达量有很好的控制,并留有浮动的空间(eg. ADH – 肝脏产生并能对抗酒精的蛋白质,越是经常饮酒肝脏里面的ADH表达量就越高,但这不是通过提高ADH基因拷贝来增加表达的,而是通过产生更多的mRNA拷贝或者更有效地利用mRNA拷贝模板来合成蛋白质)
Page 31
所有蛋白质都由基因编码,但不是所有的基因都编码蛋白质。我们的DNA中大概有20000到24000个编码蛋白质的基因。(进化让细胞通过相对少量的基因数量来创造更多种类的蛋白质)
Page 32
一条mRNA可以创造一个蛋白质,但同一条mRNA可以创造不同种的蛋白质。这是因为基因的序列中有两种不同的序列,外显子(exon,能被表达的序列),内含子(intron,不能被表达的序列),首次从DNA中拷贝出来的mRNA包含了所有的外显子和内含子,但在这之后一个多亚基蛋白质复合体会把所有的内含子序列移走并把外显子连接起来。在剪接的过程中,一部分外显子也可能被移走,或者被保留,或者被跳过,因此产生截然不同的蛋白质。细胞能够根据特定事件的需要或者接受的不同信号表达出不同的蛋白质。这也解释了为什么蛋白质往往比我们想象中的3000/3=1000个氨基酸的长度要短的原因(最常见的长度,一个基因有3000个碱基对)。
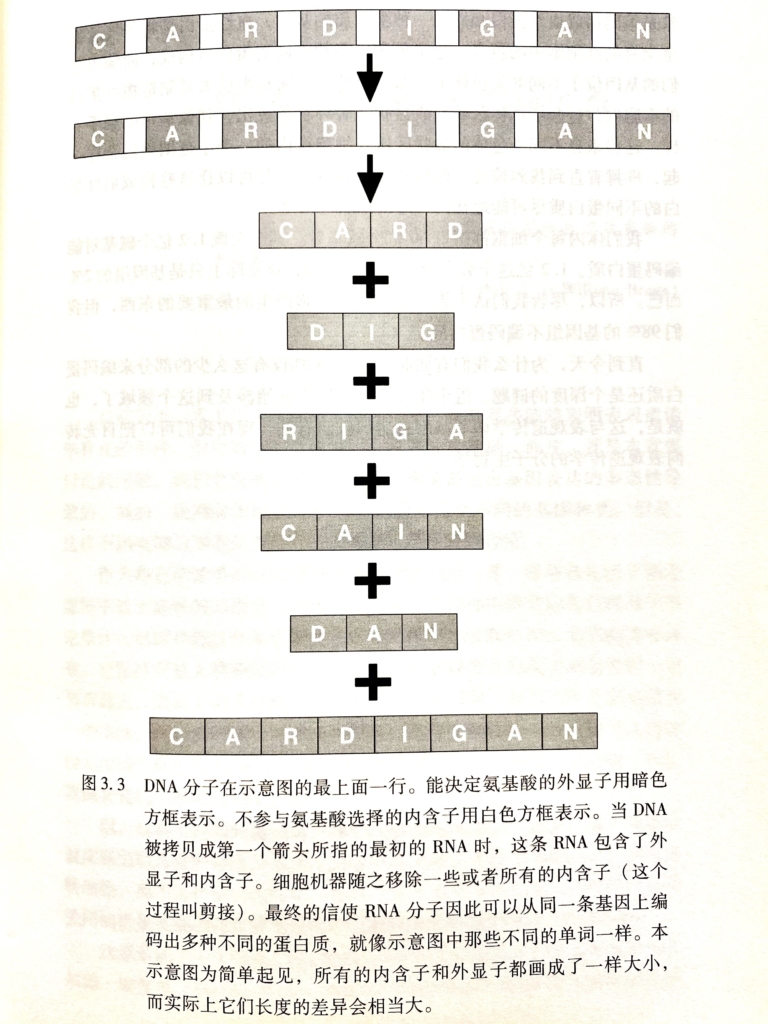
Page 33
我们体内每个细胞能携带60亿个碱基对。其中大概1.2亿个碱基对能编码蛋白质。尽管我们认为蛋白质是我们细胞产生的最重要的东西,但我们98%的基因组不编码蛋白质。
Page 35
对科学来说,探索新的思考途径比知道诸多新的事实更为重要。——Sir William Bragg
Page 36
不同的细胞具有完全相同的DNA蓝图(原作者的剧本),但携带着不同的分子修饰(拍摄剧本),并且可以在母代细胞的分裂中传递给子代细胞。简单地说,表观遗传修饰能改变一个基因表达多少,或者是否进行表达。
Page 37
DNMT(DNA MethlyTransferase)们就是一类表观遗传修饰的“书写者”——产生表观遗传编码的酶。高水平的DNA甲基化与基因关闭相关。当DNA甲基化后,它会与MeCP2(Methyl CpG binding Protein 2)的一种蛋白质相结合,这种蛋白质不会和非甲基化的CpG序列结合。MeCP2就是表观遗传编码(甲基化)的阅读者。
Page 39
MeCP2与一个基因启动子上的5-Methyl C结合后,他会结合很多其他蛋白质来帮助关闭基因,还能防止转录复合体结合到启动子,从而阻止mRNA信使分子的生成。那些启动子被高度甲基化的基因经与MeCP2结合会导致染色体该区域几乎终生关闭。该DNA会难以置信地紧密缠绕而让转录机制无法靠近碱基对来生成mRNA拷贝。(八十多年来,DNA甲基化始终保持着基因组某些区域难以置信的紧密缠绕,以保持神经元细胞相应基因的沉默。这就是我们大脑细胞不产生血红蛋白或者消化酶的原因。)
Page 39
DNMT1能够识别是否有CpG序列的甲基化仅存在于一条链中。当DNMT1发现了这种不平衡,它会把失去的甲基化加到新合成的链上。如此,子代细胞就会拥有跟母代细胞一样的甲基化特征了。

Page 41
1999年,Huda Zoghbi领导的团队证实(Rett syndrome)一种严重的自闭症(大脑发育迟滞)是由于编码甲基化DNA阅读者的基因MeCP2基因的突变造成的,由此无法生成有活性的MeCP2蛋白质,无法阅读具有完美的甲基化DNA序列。
分子表观遗传学的第二大区域——蛋白质
Page 40
组蛋白(histone)八聚体的紧凑结构由两组四种组蛋白(H2A H2B H3 H4,这些组蛋白自身折叠成紧凑的球形)构成的聚集组成,DNA则像棉花糖周围围绕的甘草一样紧紧缠绕在这个蛋白上,形成被称为核小体的结构,一个核小体上的DNA线圈包含147个碱基对。染色体的某些区域几乎始终保持着这种极端的结构,有些是一些结构性的区域,有些是被严重甲基化的区域(这也是在一些长寿细胞,如神经元细胞能够在几十年间都关闭某些基因的机制之一。)
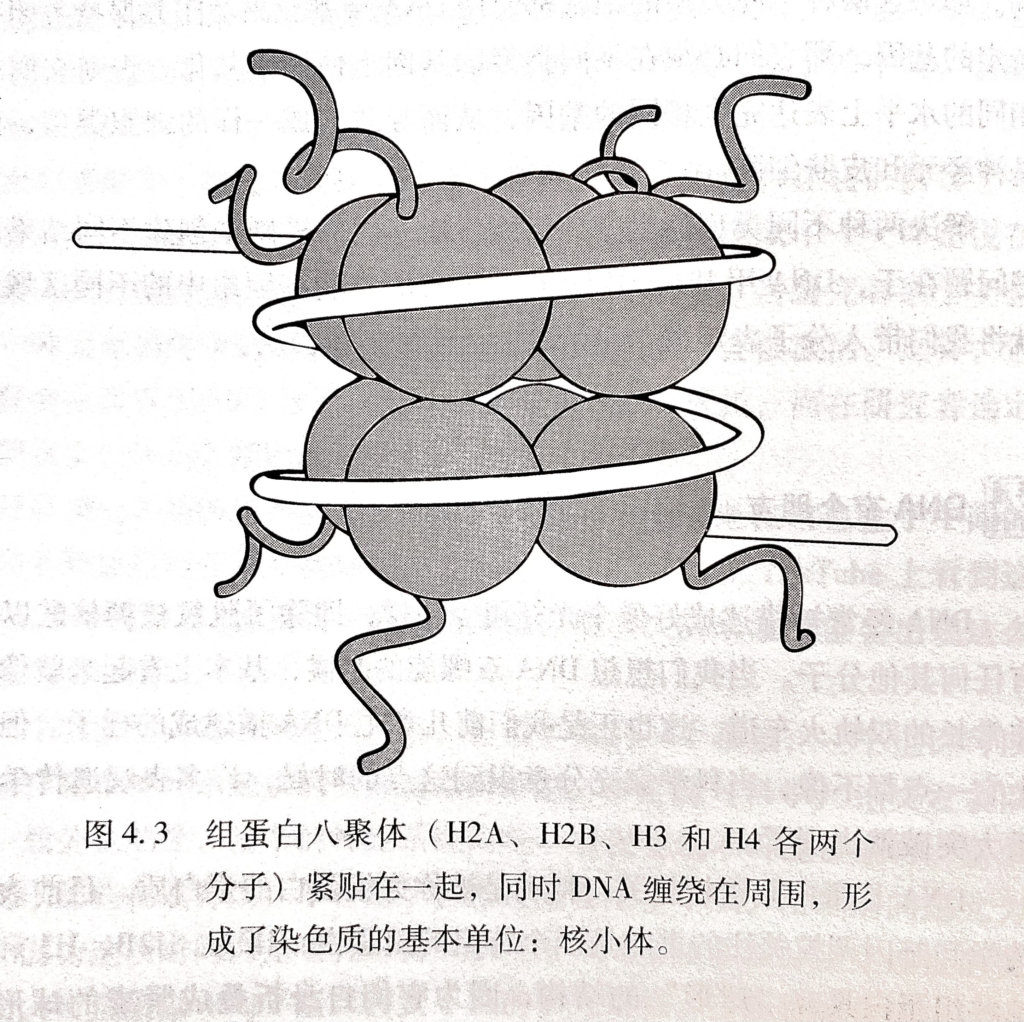
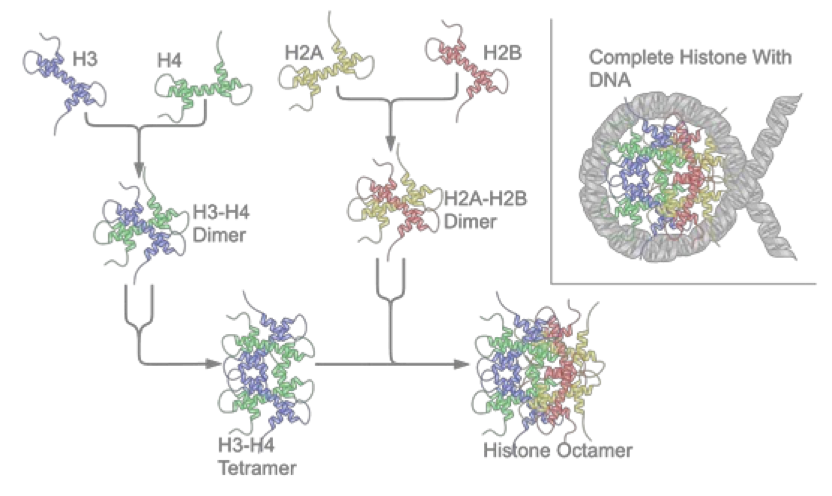

Page 41
如果DNA甲基化是《罗密欧与朱丽叶》上面的半永久标注,组蛋白上的修饰则像是铅笔做的临时批注,能挺过几次影印,但以后就会不见踪影。它们更短暂,像便利贴一样只是暂时使用。David Allis证明了细胞中的组蛋白被进行了化学修饰(+H2CO),从而增加了靠近特定修饰核小体的基因的表达。在1996年人们认为,甲基化关闭基因的表达,而组蛋白乙酰化开启基因。然而,基因很少处于全开或者全关的状态,他们更像是传统收音机上面的音量调节键。现在超过50种针对组蛋白的修饰被诸多实验室鉴定出来,有些升高基因的表达,有些则降低,这些修饰的特征被认为是一种组蛋白编码。

Page 42
但这种编码非常难读。把核小体想象成一棵装了大圣诞树的卡车,树上到处伸出的枝桠就是组蛋白尾,它们可以被表观遗传修饰来装饰。现在在不同树枝上挂上一些紫色装饰品,也可以在枝条上挂上一些绿色尖锥形状的装饰品,尽管这些树枝上已经有了紫色装饰品。同时,拿起一些红色星星,现在规定红色星星不能挂在由紫色装饰品的树枝,同样,金雪花又不能和绿色尖锥处于同一条树枝。就这样继续下去,还有更多的规矩和模式。在所有装饰品挂上去之后,我们把灯泡缠绕到了树上,每个灯泡代表一个基因。假设我们通过某一个神奇的程序根据每个灯泡周围的装饰物的精确组合来控制每个灯泡的亮度,可能很难预测大部分灯泡的亮度,因为圣诞装饰品的图案太复杂了。
Page 47
胚胎的发育就像是捕鼠器游戏。在胚胎发育最早期阶段的关键蛋白质之一就是Oct4。Oct4蛋白质能与一些关键基因结合,还可以吸引一种特定的表观遗传酶。这个酶能修饰染色质并改变基因的表达。Oct4和那个表观遗传酶都在早期胚胎发育中具有决定性作用。如果缺失任何一方,合子甚至都无法发育到ICM阶段。Oct4也能够打破已分化细胞的正常基因表达模式而使得它们变得更像干细胞,因此在正常情况下体细胞不会表达Oct4蛋白。其实早期胚胎基因表达的模式会对它们自己进行反馈,当特定的蛋白表达出来后,它们可以和Oct4启动子进行结合并关闭该基因。
Page 48
山中伸弥通过用人工的方式在已分化细胞中大量表达Oct4后,“欺骗”这些细胞具有像早期发育细胞一样行为——Oct4作为重新编程因子就连表观遗传修饰都重置了。
Page 48
表观遗传修饰在控制细胞命运方面有着重大意义。歌舞伎综合症(Kabuki syndrome)是一种复杂的发育失调,由MLL2的基因突变引起,MLL2蛋白是一种表观遗传书写者,它可以将甲基基团加到H3组蛋白第4位赖氨酸上,突变的患者不能正确书写自己的表观遗传编码。而那些移除表观遗传修饰的酶,则是表观遗传修饰的橡皮,出现突变也会导致疾病。一种PHF8的基因表达的蛋白能够移除H3组蛋白第20位赖氨酸上的甲基,它的突变会导致腭裂和身材矮小的一种综合症。
Page 49
除了书写者和橡皮,还有超过100种作为组蛋白表观遗传编码阅读者的蛋白质,这些阅读者吸引其他蛋白组成一个复合物来开启或关闭基因的表达。Eg. MeCP2帮助关闭被甲基化的DNA基因表达(见page37)
Page 49
大部分的DNA甲基化表观遗传修饰具有稳定性。而大部分组蛋白修饰具有更好的可塑性,细胞核受到任何外界刺激,作为反应都会发生这些修饰 eg.胰岛素信号,乳腺细胞对雌激素的反应…这些组蛋白编码的变化是通过后天(环境)与先天(基因)的交互以创建地球上高级生命复杂性的重要途径之一。组蛋白的修饰也允许细胞去“尝试”不同的基因表达模式,尤其在发育期。
Page 50
在发育期,当抑制性的组蛋白修饰出现在某个基因附近时,这个基因会被暂时性的失活。如果这些基因的失活对细胞有益,这些组蛋白的修饰会持续很长时间直到DNA甲基化,如果这里发生了足够多的DNA甲基化,这个基因的表达就会被关闭,而在极端环境下,整个染色体区域会变成高度紧密状态并在多次细胞分裂后仍保持失活,或者在这种不分裂的细胞中一直失活几十年。生物允许这种复杂的组蛋白修饰的方式进行微调是因为,细胞和生物需要为了适应外界环境而更好地对基因表达进行调节,eg.后天环境或者病毒的感染。
双胞胎
Page 51
同卵双胞胎=单合子双胞胎=Monozygotic。通过对一致率(concordance rate)的计算,即计算双胞胎都出现同一种疾病的比率,我们得以知道在精神分裂中遗传因素占主导地位。
Page 52
2005年, Manel Esteller领导了一个大型协作项目,而后马德里的西班牙国立癌症中心发表的一篇论文有一些有趣的发现。婴儿的MZ双胞胎染色体中无法发现组蛋白乙酰化或者DNA甲基化水平的显著差异,但老年的MZ双胞胎,比如五十多岁,却有显著差异,尤其是分开独立生活的双胞胎。这一研究结果证明了表观遗传组学(基因组上表观遗传特征的总称)是对环境差异的反应。
Page 53
即使基因完全相同的个体出生时的表观遗传学也是不同的,而这些表观遗传差异会随着年龄的增长和接触不同的环境变得更加显著。
Page 60
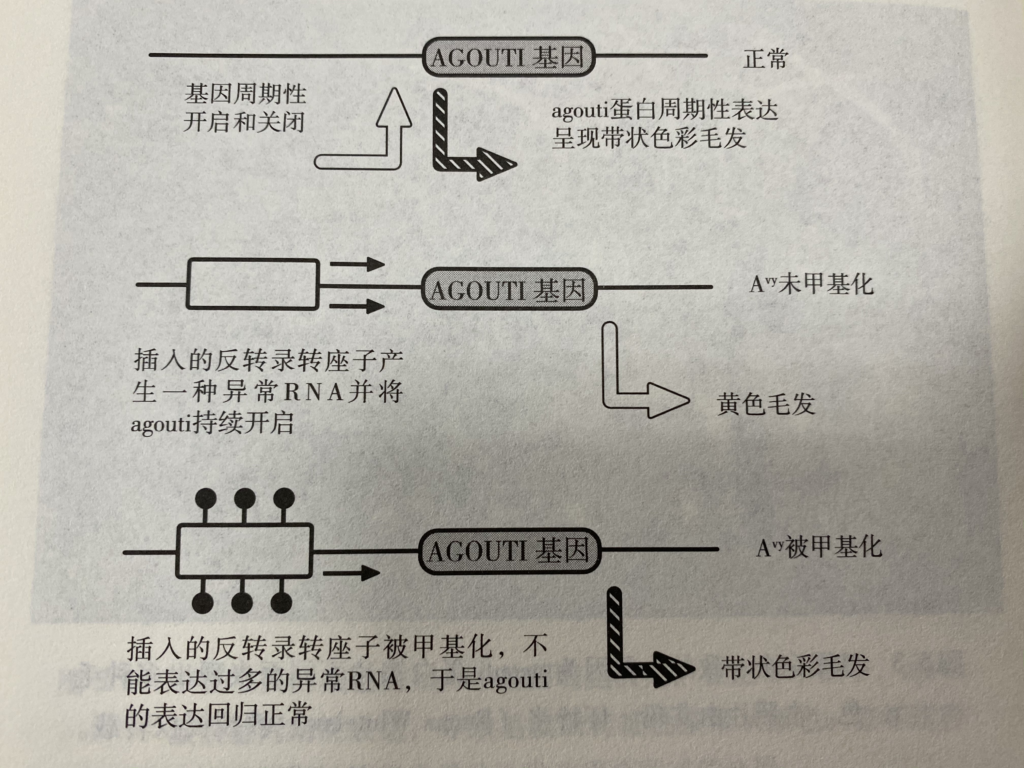
A反转录转座子DNA甲基化水平的随即多样性引起的不同毛色。发生在发育编程中的表观遗传变化的核心还是一个非特异的过程,通常被称为“随机”。
Page 62
裸露的DNA很容易被随机开启,而导致出现很多转录噪声。表观遗传蛋白有类似于减噪器的作用,它们通过用修饰来覆盖组蛋白而降低基因的表达。这是细胞中的一种复杂的平衡性,在其中表观遗传蛋白减少转录噪声而并不彻底关闭它,并通过压制的办法允许细胞有足够的基因表达灵活性来对新的信号进行反应(比如激素养分污染以及日光灯)而不会使基因随时都准备着被表达得热火朝天。发育早期是这种转录噪声控制初始建立的关键时期。(这也解释了为什么70%~80%的体重变异与后天环境、遗传无关的原因。我的理解是,表观蛋白影响基因即遗传物质表达的程度,而当某一具有相同基因的鼠群特定的表观遗传蛋白质的表达水平只有正常的一半时,它们由基因决定的特性会出现更多variance,且基因被抑制得越多,variance越大)
Page 62
DNMT3 A可以向完全未甲基化的DNA区域添加甲基基团,它负责在细胞中建立正确的DNA甲基模式,而DNMT1则是维护已经建立好的DNA甲基化模式。对于基因表达减噪工作而言,最重要的还是建立正确的DNA甲基模式(DNMT3 A)。
Page 64
组蛋白和DNA上这些最初的小幅波动,最终可能被“设置”并传递到子代细胞中。早期发育过程中表观遗传的随机波动会导致基因表达出现不完全相同的模式。
Page 65
受到(荷兰,WW2的最后一个冬天)饥荒影响而诞生的孩子,即在母亲怀孕极早期,就是发育变成处于顶峰的那个时期中,他们的细胞被表观遗传编程为尽最大努力去节约食物的模式。数据证实,通过对荷饥饿冬天幸存者进行甲基化特征分析,他们在代谢相关的关键基因上出现了变化——发育早期的低营养状态会改变代谢关键基因的表现遗传学特征。
Page 66
然而正是由于表观遗传随机多样性(随机的表观遗传波动),人群仅是患有肥胖的可能性升高,而不是“全”或“无”的类型。当然,同时针对的刺激也会改变基因的表达,例如Bisphenol A影响agouti小鼠子代的毛色分布,表明这种化学物质能够通过表观遗传机制对发育编程进行影响(Bisphenol A已经被欧盟禁止在婴儿使用的奶瓶添加)。
Page 68
进化论指出:基因的随机多样性导致了个体表型的多样化。一些个体会比其他个体更适应某些特定的环境,所以这些个体会有更多的后代。这些后代也许会遗传来自其父母的相同的优势基因型,所以它们也会成功产生更多的后代。最终,经过许多代以后,独立的种群出现了。
Page 69
拉马克提出了获得性遗传,然而收到反驳(Charles Darwin 进化论),毕竟很难想象作用于个体细胞的环境影响能激活一个特定的基因去改变碱基对序列。没有想到的是,多年以后我们发现表观遗传修饰——DNA甲基化或组蛋白修饰——确实能在细胞中通过对环境的反应而出现在特定基因上。表观遗传修饰并不能改变基因的序列,但它们确实可以调控基因表达的水平。(表观遗传修饰能从母代细胞向子代细胞传递,所以我们的眼球里才不会长出牙齿)。表观遗传改变(相对于基因来说)可以从父母向下传递给孩子,如果环境导致个体的表观遗传修饰变化也能够传递给子代,我们就会得到一个类似于拉马克遗传理论的机制。
Page 72
父亲到孩子的跨代遗传现象不是由子宫内环境或细胞质导致,表观遗传机制因此成为了解释获得性特征跨代遗传的有利候选者。
Page 74

通过小心控制宫内营养状态,对受精卵进行移植(黄色小鼠母体内的受精卵移植到深色的雌性内),以及复杂的育种计划(以此判断卵子细胞质的影响),科学家们排除了其他的一切因素并最终确定表观遗传修饰(可能是DNA甲基化)能够连同遗传密码一起被遗传。然而,控制agouti表型的DNA甲基化并不是稳定遗传的,落实到个体我们并不能精准预测。
Page 76
小鼠agouti表型和尾巴扭结表型都依赖于基因组内的特定反转录转座子,那这些到底是特例还是普遍存在的呢?首先,通过给与怀孕agouti小鼠高甲基基团饮食导致的被毛特征的变化并不能够传递给下一代。然而, 在给雄性小鼠高糖含量低蛋白的异常饮食实验中,通过对他们的后代的大量分析,研究者们发现许多参与代谢的基因的调节出现了异常。
Page 78
上述实验表明,至少在rodent animals中,父亲的饮食情况能够直接影响子代的表观遗传学修饰、基因表达和健康情况。而且这种影响出现的频繁程度让我们不能认为其成因是饮食导致的基因突变,所以,最可能的解释是饮食导致了表观遗传学影响,并可以从父母传递给孩子。(也许我们父母,甚至更上几代的祖先吃什么都会影响我们)
Page 78
题外话:广泛使用于红酒工业中的烯菌酮(Vinclozolin)一种抗真菌剂,如果被哺乳动物摄入会被转化成一种化合物并能结合在雄激素受体上——这是结合睾丸酮的受体,作为雄激素的睾丸酮对性发育、精子的生成和许多男性相关的功能非常重要。当烯菌酮与雄激素受体结合后,睾丸酮就无法将正常的信号传递给细胞,正常的激素功能也被屏蔽了。
生殖游戏
Page 79

一个革命性的概念——母本和父本的基因组可以提供相同的DNA,但他们的功能并不等效。单纯的每个基因拷贝数的正确,并不足以支持生命的正常发育和健康生活。
Page 85
葡萄胎会导致子宫中只有胎盘而没有胚胎。葡萄胎是由因不明原因丢失了细胞核的卵子受精而成的。大概80%的葡萄胎中空卵子被精子穿透而后对该精子的单倍体基因组进行拷贝并形成双倍体基因组。
Page 86
在精子穿入卵子的瞬间,雄性前核DNA上的甲基化全部以异乎寻常的速度被迅速抹去,雌性前核也在发生同样的事情,不过速度要慢很多。这意味着许多表观遗传记忆在基因组里面被剔除。抑制性DNA甲基化和组蛋白修饰被抹去的同时,其他促进基因表达的组蛋白修饰则可能被留存,因此表观遗传组正在被重新编程。
Page 86
受精卵分裂形成胚泡(ie包着高尔夫球的一个网球)——内细胞群(ICM多能细胞,可以在实验室里生长成胚胎干细胞)——ICM很快分化,借助特异性的蛋白质eg.OCT4开启另一组基因,级联表达,循环往复(参与级联表达反应的基因都是与基因组的表观遗传修饰有关的,能够改变DNA和组蛋白标记)。
Page 87
精子:特殊的染色质诱发卵子的重新编程。卵子:快速大量重新编程。卵子和精子的重新编程可以防止父母将积累的任何不适当的表观遗传修饰传递给后代。与其说擦干净黑板,倒更像是重新安装操作系统。
Page 88
亲源效应(parent-of-origin):阿奇姆·苏拉尼的研究显示我们需要雄性和雌性的前核每种都有一个才能创造出新的哺乳动物来,因为受精卵以及其分裂而来的细胞具有区分来自父亲和母亲的染色体的能力。这也产生了一个矛盾,因为这说明必定有一些表观遗传修饰能够从一代传递给下一代。
Page 89
后代的插入DNA甲基化水平依赖于其父母中传递该片段者的性别而不依赖于其父母中甲基化的程度。染色体被标记了它是从父母中哪一方来的信息。7株小鼠出现1株也说明了不是所有的基因组都携带这些信息,七分之一的出现率说明了这些标记的区域是例外,而不是常规。
Page 90
上面的第六章中,艾玛·怀特洛的工作(agouti小鼠跨代研究)向我们展示了一些表观遗传修饰确实在父母和自带中通过精子和卵子进行了传递,虽然非常罕见。所以尽管在精子和卵子融合时,绝大部分哺乳动物基因组都被进行了重置,但还是有一小部分对重新编程能够免疫。
Page 91
我们的基因组只有2%的序列编码蛋白,多达42%由反转录转座子组成,这组奇妙的DNA序列据认为是病毒整合而来。一些反转录转座子转录成为RNA并影响周围基因的表达,如引起与细胞增殖相关的基因的过度表达进而导致细胞被引向肿瘤化的道路等,造成严重的后果。针对这严重的后果,我们的细胞进化出相关机制去控制这些反转录转座子的活性使它们被甲基化,从而关闭了反转录转座子的RNA表达及对临近基因表达的干扰,其中特定一类,IAP反转录转座子是这个控制机制的特定靶点。
Page 92
在受精卵早期的重新编程过程中,我们DNA上大部分的甲基化被移除。但是IAP反转录转座子是一个例外。重新编程元件已经进化成为能够跳过这些“不法分子”而将其标记留下,是的这些反转录转座子始终保持表观遗传抑制状态,降低风险。巧合的是,在研究非基因特征跨代遗传的两种模型(agouti和Axin^Fu小鼠)中的表型都是由特定基因上游的IAP反转录转座子的甲基化水平而决定的,父母的DNA甲基化水平传递给下一代,并由反转录转座子的表达水平决定表型如何。
Page 91
一些上文提到过的跨代遗传的例子,包括营养对后代的影响,烯菌酮的跨代作用等,一些环境刺激导致的配子染色体的表观遗传学改变,可能存在于那些在精子和卵子融合后立刻进行的重新编程的特赦区域内。
第八章 性别战争
Page 93


